D
DesertEagle
Такое ощущение, что многие махолетчики живут в 16 веке, когда еще не было изобретено фотоаппаратов, не говоря уж про базовые понятия из аэродинамики.
Что, так сложно найти видео полета какой-нибудь птицы, сделать оттуда несколько кадров да самому посмотреть, раз уж не верите тому что пишут умные люди в книгах про птиц, начиная с полутора столетий назад (с Отто Лилиенталя)?
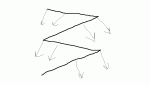
Длины стрелок для двух сечений, примерно в середине крыла и ближе к кончику, я нарисовал от балды, но думаю принцип должен быть понятен. Синяя стрелка вперед у обоих сечений одинаковая, так как это скорость полета птицы. Зеленая на кончике длиннее, так как окружная скорость кончиков выше.
Реальная скорость движения сечения крыла относительно воздуха получается по голубой стрелке. Именно это направление набегающего потока, и именно относительно него строятся углы атаки. А теперь смотрим под каким углом к потоку идет каждое из этих двух сечений и делаем выводы.
А теперь вопрос персонально к KarpovAndrey, ну и чему получились равны углы атаки, отмеченные красным, в конкретно этот момент маха? Ну там близко к 10 град, а может 90 град, а может -20 град? А теперь домашнее задание: взять реальные значения скорости полета конкретной птицы и ее частоту махов. И аккуратно нарисовать такие вектора, соблюдая их реальные длины, для каждого сечения в каждый момент маха.
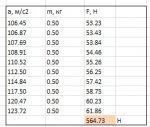
И, о чудеса науки, оказывается, можно не просто их рисовать, а еще и по формулам аэродинамики найти длины стрелок подъемной силы и лобового сопротивления, которые привязаны к этим векторам воздушных скоростей. И определить на их основе создаваемую птицей подъемную силу конкретно в граммах/килограммах. Да, я понимаю, что для большинства махолетчиков это открытие сейчас было как гром среди белого дня, но что делать, с тех пор как вы остановились в развитии, наука ушла далеко вперед. Более того, оказывается можно сравнить полученный результат с известным весом птицы. Проверив таким образом правильность расчета. А то мало ли, вдруг формулы аэродинамики не подходят для живой птицы? Обязательно надо сравнить. После этого можете убрать из формул движение вперед-назад (вашу любимую восьмерку) и сделать расчет чисто для движения крыла вверх-вниз. И сравнить результаты, выявив таким образом вклад восьмерки в общие аэродинамические силы при машущем полете. И удивиться.
Более того, эти же стрелки и скорость их перемещения, нарисованные прямо на птице из любого видеоролика, однозначно определяют мускульную мощность, затрачиваемую птицей на махи и на полет. Пожалуйста, махолетчики со слабым здоровьем, не делайте таких расчетов. Потому что я представляю какой для вас будет удар полученные цифры. Еще бы, столько лет удивляться эффективности птичьего полета, читая рассказы специалистов, которые одинаково специалисты как в машущем полете, так и в пользе шапочек из фольги для защиты от излучения американских спецслужб; травить умильные байки с товарищами про 1.8 Вт затрачиваемой мощности в полете у 12 кг гуся, охать и ахать, мечтая познать тонкости нестационарной аэродинамики, дающей птицам такие потрясающие характеристики. А тут любой видеоролик из интернета, карандаш и знания из второго класса (или когда там школьники начинают проходить про синусы и косинусы?), оказывается, все расставляют по местам. Понимаю, не каждый выдержит такое потрясение. Так что пожалуйста махолетчики, не надо так делать. Не рисуйте никаких векторов на птицах и не делайте никаких расчетов, поберегите себя.
Что, так сложно найти видео полета какой-нибудь птицы, сделать оттуда несколько кадров да самому посмотреть, раз уж не верите тому что пишут умные люди в книгах про птиц, начиная с полутора столетий назад (с Отто Лилиенталя)?
Длины стрелок для двух сечений, примерно в середине крыла и ближе к кончику, я нарисовал от балды, но думаю принцип должен быть понятен. Синяя стрелка вперед у обоих сечений одинаковая, так как это скорость полета птицы. Зеленая на кончике длиннее, так как окружная скорость кончиков выше.
Реальная скорость движения сечения крыла относительно воздуха получается по голубой стрелке. Именно это направление набегающего потока, и именно относительно него строятся углы атаки. А теперь смотрим под каким углом к потоку идет каждое из этих двух сечений и делаем выводы.
А теперь вопрос персонально к KarpovAndrey, ну и чему получились равны углы атаки, отмеченные красным, в конкретно этот момент маха? Ну там близко к 10 град, а может 90 град, а может -20 град? А теперь домашнее задание: взять реальные значения скорости полета конкретной птицы и ее частоту махов. И аккуратно нарисовать такие вектора, соблюдая их реальные длины, для каждого сечения в каждый момент маха.
И, о чудеса науки, оказывается, можно не просто их рисовать, а еще и по формулам аэродинамики найти длины стрелок подъемной силы и лобового сопротивления, которые привязаны к этим векторам воздушных скоростей. И определить на их основе создаваемую птицей подъемную силу конкретно в граммах/килограммах. Да, я понимаю, что для большинства махолетчиков это открытие сейчас было как гром среди белого дня, но что делать, с тех пор как вы остановились в развитии, наука ушла далеко вперед. Более того, оказывается можно сравнить полученный результат с известным весом птицы. Проверив таким образом правильность расчета. А то мало ли, вдруг формулы аэродинамики не подходят для живой птицы? Обязательно надо сравнить. После этого можете убрать из формул движение вперед-назад (вашу любимую восьмерку) и сделать расчет чисто для движения крыла вверх-вниз. И сравнить результаты, выявив таким образом вклад восьмерки в общие аэродинамические силы при машущем полете. И удивиться.
Более того, эти же стрелки и скорость их перемещения, нарисованные прямо на птице из любого видеоролика, однозначно определяют мускульную мощность, затрачиваемую птицей на махи и на полет. Пожалуйста, махолетчики со слабым здоровьем, не делайте таких расчетов. Потому что я представляю какой для вас будет удар полученные цифры. Еще бы, столько лет удивляться эффективности птичьего полета, читая рассказы специалистов, которые одинаково специалисты как в машущем полете, так и в пользе шапочек из фольги для защиты от излучения американских спецслужб; травить умильные байки с товарищами про 1.8 Вт затрачиваемой мощности в полете у 12 кг гуся, охать и ахать, мечтая познать тонкости нестационарной аэродинамики, дающей птицам такие потрясающие характеристики. А тут любой видеоролик из интернета, карандаш и знания из второго класса (или когда там школьники начинают проходить про синусы и косинусы?), оказывается, все расставляют по местам. Понимаю, не каждый выдержит такое потрясение. Так что пожалуйста махолетчики, не надо так делать. Не рисуйте никаких векторов на птицах и не делайте никаких расчетов, поберегите себя.
Вложения
-
 17,6 КБ Просмотры: 109
17,6 КБ Просмотры: 109








")


