D
DesertEagle
Читаем дальше... Теперь выдержки из документа: The vortex wake of blackcaps (Sylvia atricapilla L.) measured using high-speed
digital particle image velocimetry (DPIV).
Эта работа была проведена после предыдущих DPIV видеосъемок стрижей, малиновок и летучших мышей (см. переводы обзоры на пред.страницах), поэтому в ней упор был сделан на съемку области ближе к крыльям, в попытке объяснить разницу в вихревом следе у летучих мышей и у птиц. Одной из версий было предположение, что у птиц хвост помогает организовать вихри у корневых частех крыла. Летучие мыши все безхвостые, а некоторые и с "отрицательным" хвостом, то есть выемкой между задних лап. Для этого у одной из испытуемых птиц был полностью обрезан хвост, а вторая оставлена с хвостом.
Исследовались птицы черноголовки:
Масса: 16.3 (без хвоста) и 19.5 грамм (с хвостом)
Площадь крыльев: 0.0111 и 0.0105 м2
Скорость полета: 21.6 - 36 км/час (наиболее экономичная 23-24 км/час)
Удлинение: 5.1 и 5.2
Частота махов: 17-18 Гц
Re: 17500-29500
Съемка хвостатых и безхвостых птиц не выявила разницы в вихревом следе. Откуда можно сделать вывод, что хвост птицы не участвует в процессе машущего полета.
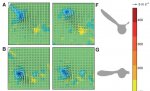
Когда начинается мах вниз, образуется мощный стартовый вихрь за задней кромкой крыла. Подъемная сила и тяга достигают максимума к 40% маха вниз, после чего постепенно ослабляются до окончания маха вниз и далее продолжают ослабляться в течении всего маха вверх, пока в конце маха вверх не обратятся в ноль. Причем сопротивление при махе вверх падает быстрее, чем подъемная сила. Разницы между взаимодействием концевых и корневых вихрей у хвостатой и безхвостой птицы не выявлено. Таким образом, гипотеза о каком либо влиянии на махи хвоста у птиц по сравнению с летучими мышами, не подтвердилась.
При махе вверх создается подъемная сила и лобовое сопротивление, но меньшие по величине, чем при махе вниз. Тяга при махе вверх не создается.
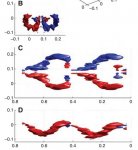
Как и у других птиц и у летучих мышей, у черноголовок существует объединение и разъединение малозначительных вихрей крыльев и корпуса в разные моменты махов.
Точность измерения в этом эксперименты была довольно высока: найденная по вихревому следу подъемная сила составила 99.7% от веса птицы для безхвостой птицы и 105.2% для хвостатой. То есть ошибка замеров составила не более 0.3-5.2%.
Средний Cy составил 1.3 на минимальной скорости полета и 0.16 на максимальной.
Средний Cx составил 0.21 на минимальной скорости полета и 0.051 на максимальной.
Таким образом, аэродинамическое качество получилось 6.19 на скорости 21.6 км/час и 3.14 на скорости 36 км/час. Максимальное аэродинамическое качество получилось 7.6 единиц на наиболее экономичной скорости (сюрприз, да) полета 23 км/час.
Причем у безхвостой птицы качество оказалось заметно выше, почти на единицу во всех режимах полета. Таким образом, хвост у птицы в полете создает исключительно сопротивление, но нужен для маневренности и снижения скорости при посадке. А в полете птицы хвост убирают/сжимают в одну линию, чтобы он меньше мешался. Хотя это можно списать на разницу в летном стиле двух разных испытуемых птиц, но другие исследования по сопротивлению чучел птиц с хвостом и без, подтверждают увеличение лобового сопротивления при удлинении хвоста, что сходится с результатами этого исследования.
Не найдено подтверждения гипотезе, что хвост может утилизировать вихри, сходящие с корневых частей крыльев (как в эффекте машущего предкрылка Болдырева с неподвижным основным крылом за предкрылком) и тем самым повышать летные свойства птиц. Реальное течение воздушного потока в окрестностях хвоста не совпадает с предсказываемым такой гипотезой.
Также упоминается о другом эксперименте, где исследовался один соловей сначала с хвостом, а потом он же без хвоста. На вихревом следе ничего принципиально не изменилось.
Нестационарных эффектов при махах не обнаружено. Полет черноголовок в целом похож на быстрый полет соловья. Общего с летучими мышами только небольшой противоположный вихрь на корпусе в начале маха, который потом пропадает/объединяется с крыльевыми. Это та часть, которая обсуждалась об объединении крыльев с корпусом в единое "крыло" в определенном момент маха, когда говорили о мышах. Только у черноголовок это выражено намного слабее. Это говорит о том, что по крайней мере частично в начальный момент махов крылья черноголовки работают по отдельности, или другими словами, корпус не участвует в создании аэродинамических сил при махе вниз (просто имеет минимально возможное лобовое сопротивление и все). Другие исследования показывают, что корпус птицы начинает играть значимую роль в создании аэродинамических сил только при прерывистом полете с падениями со сложенными крыльями. Там корпус создает положительную подъемную силу, является несущим корпусом, позволяющем такому падающему воробью с убранными крыльями иметь аэродинамическое качество до 1.4 единиц. А в обычном машущем полете корпус птицы создает только минимальное лобовое сопротивление.
digital particle image velocimetry (DPIV).
Эта работа была проведена после предыдущих DPIV видеосъемок стрижей, малиновок и летучших мышей (см. переводы обзоры на пред.страницах), поэтому в ней упор был сделан на съемку области ближе к крыльям, в попытке объяснить разницу в вихревом следе у летучих мышей и у птиц. Одной из версий было предположение, что у птиц хвост помогает организовать вихри у корневых частех крыла. Летучие мыши все безхвостые, а некоторые и с "отрицательным" хвостом, то есть выемкой между задних лап. Для этого у одной из испытуемых птиц был полностью обрезан хвост, а вторая оставлена с хвостом.
Исследовались птицы черноголовки:
Масса: 16.3 (без хвоста) и 19.5 грамм (с хвостом)
Площадь крыльев: 0.0111 и 0.0105 м2
Скорость полета: 21.6 - 36 км/час (наиболее экономичная 23-24 км/час)
Удлинение: 5.1 и 5.2
Частота махов: 17-18 Гц
Re: 17500-29500
Съемка хвостатых и безхвостых птиц не выявила разницы в вихревом следе. Откуда можно сделать вывод, что хвост птицы не участвует в процессе машущего полета.
Когда начинается мах вниз, образуется мощный стартовый вихрь за задней кромкой крыла. Подъемная сила и тяга достигают максимума к 40% маха вниз, после чего постепенно ослабляются до окончания маха вниз и далее продолжают ослабляться в течении всего маха вверх, пока в конце маха вверх не обратятся в ноль. Причем сопротивление при махе вверх падает быстрее, чем подъемная сила. Разницы между взаимодействием концевых и корневых вихрей у хвостатой и безхвостой птицы не выявлено. Таким образом, гипотеза о каком либо влиянии на махи хвоста у птиц по сравнению с летучими мышами, не подтвердилась.
При махе вверх создается подъемная сила и лобовое сопротивление, но меньшие по величине, чем при махе вниз. Тяга при махе вверх не создается.
Как и у других птиц и у летучих мышей, у черноголовок существует объединение и разъединение малозначительных вихрей крыльев и корпуса в разные моменты махов.
Точность измерения в этом эксперименты была довольно высока: найденная по вихревому следу подъемная сила составила 99.7% от веса птицы для безхвостой птицы и 105.2% для хвостатой. То есть ошибка замеров составила не более 0.3-5.2%.
Средний Cy составил 1.3 на минимальной скорости полета и 0.16 на максимальной.
Средний Cx составил 0.21 на минимальной скорости полета и 0.051 на максимальной.
Таким образом, аэродинамическое качество получилось 6.19 на скорости 21.6 км/час и 3.14 на скорости 36 км/час. Максимальное аэродинамическое качество получилось 7.6 единиц на наиболее экономичной скорости (сюрприз, да) полета 23 км/час.
Причем у безхвостой птицы качество оказалось заметно выше, почти на единицу во всех режимах полета. Таким образом, хвост у птицы в полете создает исключительно сопротивление, но нужен для маневренности и снижения скорости при посадке. А в полете птицы хвост убирают/сжимают в одну линию, чтобы он меньше мешался. Хотя это можно списать на разницу в летном стиле двух разных испытуемых птиц, но другие исследования по сопротивлению чучел птиц с хвостом и без, подтверждают увеличение лобового сопротивления при удлинении хвоста, что сходится с результатами этого исследования.
Не найдено подтверждения гипотезе, что хвост может утилизировать вихри, сходящие с корневых частей крыльев (как в эффекте машущего предкрылка Болдырева с неподвижным основным крылом за предкрылком) и тем самым повышать летные свойства птиц. Реальное течение воздушного потока в окрестностях хвоста не совпадает с предсказываемым такой гипотезой.
Также упоминается о другом эксперименте, где исследовался один соловей сначала с хвостом, а потом он же без хвоста. На вихревом следе ничего принципиально не изменилось.
Нестационарных эффектов при махах не обнаружено. Полет черноголовок в целом похож на быстрый полет соловья. Общего с летучими мышами только небольшой противоположный вихрь на корпусе в начале маха, который потом пропадает/объединяется с крыльевыми. Это та часть, которая обсуждалась об объединении крыльев с корпусом в единое "крыло" в определенном момент маха, когда говорили о мышах. Только у черноголовок это выражено намного слабее. Это говорит о том, что по крайней мере частично в начальный момент махов крылья черноголовки работают по отдельности, или другими словами, корпус не участвует в создании аэродинамических сил при махе вниз (просто имеет минимально возможное лобовое сопротивление и все). Другие исследования показывают, что корпус птицы начинает играть значимую роль в создании аэродинамических сил только при прерывистом полете с падениями со сложенными крыльями. Там корпус создает положительную подъемную силу, является несущим корпусом, позволяющем такому падающему воробью с убранными крыльями иметь аэродинамическое качество до 1.4 единиц. А в обычном машущем полете корпус птицы создает только минимальное лобовое сопротивление.